植物のCO2センシングの分子機構
 近年、大気中の CO2 濃度の急激な増加による地球温暖化が問題となっており、今世紀後半には産業革命以前の約2倍の CO2 濃度になると予測されています。植物は CO2を吸収し光合成を行うので、これからの地球環境問題を考える上で植物の CO2感知の機構を解明することは重要な課題です。CO2は植物の表面にある気孔という穴から取り込まれますが、この穴はCO2濃度によって開度が変化し、低CO2条件下で開き、高 CO2条件下で閉じます。このようなCO2による気孔開閉の応答のメカニズムを解明するために、私たちの研究室では遺伝学的手法を取り入れた研究を行っています。 近年、大気中の CO2 濃度の急激な増加による地球温暖化が問題となっており、今世紀後半には産業革命以前の約2倍の CO2 濃度になると予測されています。植物は CO2を吸収し光合成を行うので、これからの地球環境問題を考える上で植物の CO2感知の機構を解明することは重要な課題です。CO2は植物の表面にある気孔という穴から取り込まれますが、この穴はCO2濃度によって開度が変化し、低CO2条件下で開き、高 CO2条件下で閉じます。このようなCO2による気孔開閉の応答のメカニズムを解明するために、私たちの研究室では遺伝学的手法を取り入れた研究を行っています。
気孔は開けば開くほど蒸散(植物体内からの水分の蒸発)が盛んになり、気化熱が奪われるため葉面温度は低下します(これはヒトが汗をかくと体温が下がるのと同じ原理です)。私たちは気孔の開度の変化を葉の表面の温度でモニターし、CO2応答に異常をもつシロイヌナズナの変異株をスクリーニングすることに成功しました ( Hashimoto et al., Nature Cell Biol. 2006; Negi et al., Nature 2008;総説として 祢宜 淳太郎他 蛋白質 核酸 酵素 54(6),707-715, 2009 を参照)。
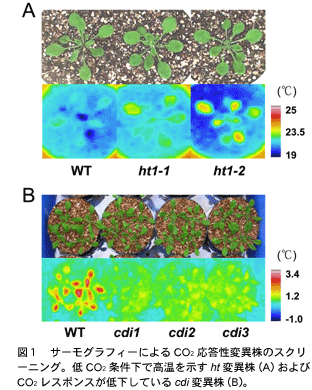
サーモグラフィーを用いたサーマルイメージングによる突然変異体の単離例を図1に示しています。Aでは葉面温度(絶対温度)を指標にしたスクリーニングによって単離された ht1 (high leaf temperature 1) 変異体の画像を示しています。ht1-1 およびそのアリルの ht1-2 は低CO2条件 (100 ppm) でも葉温が野生株(WT)と比べると高温であることがわかります。Bでは葉面温度変化(サブトラクション画像)を指標にしたスクリーニング、つまり、低 CO2条件のときの画像と高CO2条件 (700 ppm) のときの画像の差を指標にして単離された cdi(carbon dioxide insensitive)変異体を示しています。WT では CO2変化に伴う葉温変化が大きいのに対し、3つの cdi 変異株 (cdi1-cdi3) では変化が小さいことがわかります。
これらの変異体のうち、ht1 変異体では低 CO2条件下においても葉面温度が高く、CO2濃度を変化させたときの葉面温度の変化が小さいことがわかりました。これは、気孔の開度が低く、CO2濃度を変化させても気孔開度があまり変化しないことに起因します。また、ht1変異体の一つであるht1-2株では CO2 応答性が完全に消失しているのに対し、CO2以
外の環境刺激である青色光やアブシジン酸(気孔を閉じさせる働きがある植物ホルモン)による応答性は損なわれません。この変異体の原因遺伝子をクローニン
グして解析した結果、キナーゼ(タンパク質リン酸化酵素)をコードしており、孔辺細胞(気孔を形作る細胞)で特異的に働くことが分かりました。また、図2
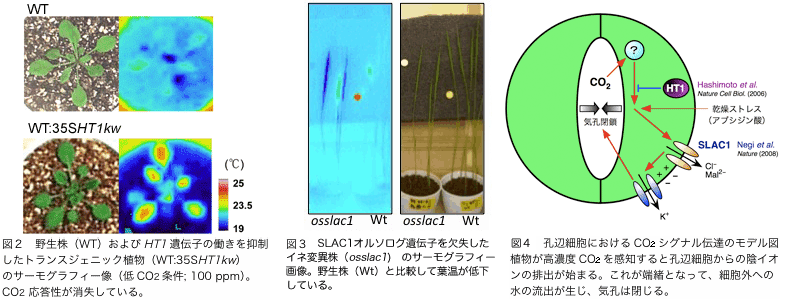
に示すとおり、人為的にキナーゼ活性を消失するように設計したトランスジェニック植物(ドミナントネガティブ形質転換植物;WT::35SHT1kw)で
はht1-2株と同様、低 CO2条件下でも高温を示し、CO2応答性が消失しました。これらの実験から、HT1 キナーゼは植物におけるCO2シグナル伝達系に重要な役割を担っていると考えられます。
一方、画像の差を指標にして同定された cdi3変異の原因遺伝子は、最近の研究から、気孔の細胞膜に局在する排出型有機酸トランスポーター/陰イオンチャネルタンパク質であることが分かってきました(遺伝子の名前もCDI3からSLAC1, SLOW ANION CHANNEL 1と改名)。このタンパク質は乾燥、CO2 などの外部環境の変化に伴う気孔閉鎖に必須の因子であると考えられていましたが、これまでその分子実体は不明でした。このSLAC1遺伝子については、イネを材料としてオルソログ遺伝子を機能欠失した突然変異株の解析も進めています。同変異株においても、cdi3と同様に気孔の開口や葉面温度の低下が見られることから(図3)、CO2取り込み能や高温耐性能を付加するツールとしての利用が期待できます。
図4に、孔辺細胞のCO2シグナル伝達経路におけるHT1とSLAC1の位置について示しました。現在、新たなCO2シグナル関連因子の同定をこのような遺伝学的アプローチによって進めており、植物におけるCO2感知・適応のメカニズムを分子レベルで解明することを目指しています。
|
 English
English 




